受体 (生物化学)

圖爲受體作用機理的示意簡圖。細胞外的配體分子(圖中標爲1)在與受體(圖中標爲2)結合後,受體結構發生改變,使細胞內產生相應的效應,這可能是由第二信使分子(圖中標爲3)誘導的
受體(Receptor),又称受器、接收器,是一個生物化學上的概念,指一類能傳導細胞外信號,並在細胞內產生特定效應的分子。產生的效應可能僅在短時間內持續,比如改變細胞的代謝或者細胞的運動。也可能是長效的效應,比如上調或下調某個或某些基因的表達[1][2][3]。
受體通過與特定的配體結合而感知到細胞外的信號。隨後,受體的結構發生變化,並誘導細胞內產生相應的效應。受體通過信号级联效應,逐步以指數級擴大細胞內產生的效應的強度。信號級聯的第一步可能是產生cAMP等第二信使分子,誘導下一級反應[3]。根據受體所在的位置,可以分爲細胞表面受體和細胞內受體兩類。其中細胞表面受體位於細胞表面,處於內環境中的配體可以直接與之結合。大部分的細胞內受體都屬於核受體。在未與配體結合時,這些受體位於細胞質中,配體需要穿過細胞膜進入細胞內,才能與該受體結合結合。在與配體結合後,核受體會轉入細胞核中發揮效應。另一類細胞內受體是細胞內的酶、RNA、核糖體等,配體通過與這些受體結合發揮效應[4][5]。
目录
1 細胞表面受體
1.1 G蛋白偶聯受體
1.2 受體酪氨酸激酶
1.3 鳥苷酸環化酶偶聯受體
1.4 離子通道
1.5 黏附受體
2 細胞內受體
3 信號級聯
4 受體脫敏
5 学说发展史
6 參見
7 參考
細胞表面受體
一種G蛋白偶聯受體的結構示意圖,圖中用彩色標出的是其七次穿膜的結構
細胞表面受體是一類位於細胞表面的受體,可與細胞外的配體結合結合,並引發細胞內的反應。目前已發現的細胞表面受體中,較經典的有5類:G蛋白偶聯受體(G protein-coupled receptor)、受體酪氨酸激酶(Receptor tyrosine kinases)、鳥苷酸環化酶偶聯受體(Guanylate cyclase-coupled receptor)、離子通道(ion channel),以及黏附受體(adhesion receptor),其中以G蛋白偶連受體的數量最多,相關的研究也是最深入的,諾貝爾化學獎以及諾貝爾生理醫學獎都有授予給G蛋白偶聯受體的研究者的紀錄[6][7][3][8]。
G蛋白偶聯受體
G蛋白偶聯受體是目前發現最主要的一種細胞表面受體,對它的研究也是最透徹的。G蛋白偶聯受體可分爲A-F六類,不同分類之間並沒有顯著的同源關係[9][10][11][12]。G蛋白偶聯受體的一個特徵是,它有一個穿膜7次結構域的受體,肽鏈的N端位於細胞外側,C端則位於細胞內。另一個特徵則是該受體與G蛋白偶聯。G蛋白通常含有三個亞基α、β、γ,其中α亞基靜息狀態下與GDP結合。當受體被配體激活後,受體會使與G蛋白結合的GDP轉化爲GTP,從而使G蛋白活化,α亞基分離,引起下一步反應,通過肌醇三磷酸(IP3)、cAMP等第二信使引發下一級反應[3][8]。
受體酪氨酸激酶
受體酪氨酸激酶是一類有著單次穿膜結構域的受體,在細胞內側偶聯着酪氨酸激酶。當該類受體激活後,內側的酪氨酸激酶會發生活化,對蛋白肽鏈上的酪氨酸(Tyr)進行磷酸化,進而產生下一步效應。胰島素受體、IGF1R,以及IGF2R都屬於受體酪氨酸激酶[13][14][8]。
鳥苷酸環化酶偶聯受體
鳥苷酸環化酶偶聯受體(Guanylate cyclase-coupled receptor),亦稱爲受體鳥苷酸環化酶(Receptor guanylate cyclase),是一種單次穿膜的細胞表面受體,該受體因爲可在激活後將鳥苷酸環化爲cGMP,以cGMP分子作爲第二信使而得名。在人體內,因爲不同組織中鳥苷酸環化酶偶聯受體的類型不同,這類受體的具體功能也不同。腎臟中的心房利鈉肽(ANF)受體NRP1、NRP2、NRP3就屬於鳥苷酸環化酶偶聯受體。另外,亦存在位於細胞內的鳥苷酸環化酶偶聯受體,比如可溶性一氧化氮激活鳥苷酸環化酶(soluble NO-activated guanylate cyclase)[8][15]。
離子通道
離子通道是一類位於細胞表面的蛋白,平時屬於關閉狀態,離子不能通過,在被激活後,通道才會打開,離子方可通過離子通道轉移到細胞膜另一側。一類離子通道,即配體門控離子通道(Ligand-gated ion channel)可在與配體結合後激活,使對應的離子得以通過。神經突觸中突觸後膜上的毒蕈鹼受體就是一類配體門控離子通道,在與乙醯膽鹼結合後,會打開離子通道,允許鈣離子(Ca2+)、鈉離子(Na+)內流,引發突觸後膜興奮[16][17][18]。
黏附受體
黏附受體(adhesion receptor)是一類與細胞黏附相關的受體,主要接收與細胞黏附的信號。細胞膜上的整合素就是一種黏附受體。整合素在細胞外會與配體膠原蛋白、硫酸乙酰肝素等結合,而細胞內的結構域則是與細胞骨架結合(結合可能是直接結合,也可能由talin蛋白等作爲連接的中間體),通過感知細胞外膠原蛋白等的變化,調節細胞內的細胞極性、細胞生長存活狀態,以及基因表達強度等,同時,整合素也可以反過來對細胞外產生影響,如調節細胞黏附和遷移,以及細胞外基質的聚合[8]。
細胞內受體

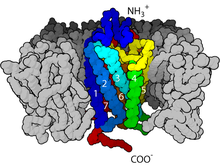
圖爲一種核受體PPAR-γ與DNA結合結構的晶體衍射圖
位於細胞內的受體稱爲細胞內受體。大部分的細胞內受體是核受體,在未激活時位於細胞質中。因爲配體需要穿過細胞膜,擴散入細胞質中後才可以與核受體結合,因而核受體的配體大部分是小分子脂類物質。核受體在與配體結合後,會進入細胞核內,對基因的表達進行調控,因而核受體可認爲是一類轉錄因子。因爲核受體的效應是改變基因表達,因而相比細胞表面受體,核受體產生效應所需時間相對較長,同時產生的效應相對也能持續較長時間[19][20][5]。
除核受體外,還有一類細胞內受體的化學本質是細胞內的酶、結構性蛋白、核糖體或RNA。比如藥物(配體)甲氧苄氨嘧啶可以作用於受體微管、紅黴素可以作用於細菌的50s核糖體亞基[5]。
信號級聯
受體在接受胞外信號後,會向細胞內傳遞信號。在這個過程中,信號通過不斷向下一級傳遞的方式逐漸增大,這個過程稱爲信號級聯。雖然受體最初只能產生小部分的效應分子,但每個效應分子又能激活幾個下一級的信號分子,這樣,一個受體的信號會以指數級的方式不斷擴大,最後在細胞內產生明顯的效應,比如使數個基因的表達發生變化。一些受體在激活後會產生一些效應分子,如cAMP等,並由這些分子開始信號級聯反應,這樣的分子稱爲第二信使[21][22][8]。
受體脫敏

受體脫敏的圖解。可見受體在反覆受到配體刺激後,可通過內吞作用被運回細胞內部,使配體無法與之結合
受體脫敏(desensitization)是指受體在反覆受到配體刺激活化後,對配體反應的敏感性的強度下降的現象。在藥理學中,快速抗藥即由受體脫敏造成。受體脫敏的一種機制是,受體通過內吞作用被運回細胞內部,使細胞表面的受體數量減少,能與內環境中配體結合的受體也就減少。此外,一部分受體,如一些離子通道,在激活後需要經過一定的靜息期方可再次被激活[5][8]。
学说发展史
1878年,朗格莱(Langley)提出了最早的受体假设,他用“受体物质”来解释阿托品和毛果芸香碱对猫唾液分泌的拮抗作用。1913年,欧利希(Ehrlich)根据实验结果提出了“锁和钥匙”的药物与受体的互补关系,但当时认为受体和配体都是静止不动的。1933年,克拉克(Clark)在研究药物对蛙心的量效关系中,定量的阐明了药物于受体的相互作用。这些说法为受体学说奠定了基础。[23]
20世纪50-60年代,Ariens和斯蒂芬森(Stephenson)从实验发现药物产生最大效应不一定占领全部受体,由此提出的备用受体学说和速率学说,从动力学的角度解释了受体拮抗剂和激动剂的作用。[23]
20世纪70年代以后,随着蛋白质晶体学的发展,许多配体和受体的结构被人们所认识,从而阐明了受体亚型、离子通道等的分布和功能。变构学说彻底打破了蛋白质静止不动的认识,认为受体在有活性与无活性的构象状态间转化。1977年,格里夫斯(Greaves)提出的能动受体学说,把受体的微观变化同生理、生化或药理反应相联系,说明了受体在细胞膜内传递信息的作用机制。[23]
參見
- 配體 (生物化學)
受體拮抗劑和激動劑
參考
^ Hall, JE. Guyton and Hall Textbook of Medical Physiology. Philadelphia, PA: Elsevier Saunders. 2016: 930–937. ISBN 978-1-4557-7005-2.
^ Alberts, Bruce; Bray, Dennis; Hopkin, Karen; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walter, Peter. Essential Cell Biology Fourth. New York, NY, USA: Garland Science. 2014: 534. ISBN 978-0-8153-4454-4. 使用|accessdate=需要含有|url=(帮助)
^ 3.03.13.23.3 Harvey Lodish; 等. Chapter 15. Molecular Cell Biology (7th edition). Macmillan Higher Education. 2013: 674–683. ISBN 978-1-4641-0981-2. 引文格式1维护:显式使用等标签 (link)
^ Rang HP, Dale MM, Ritter JM, Flower RJ, Henderson G. Rang & Dale's Pharmacology 7th. Elsevier Churchill Livingstone. 2012. ISBN 978-0-7020-3471-8.
^ 5.05.15.25.3 Michelle A. Clark; 等. Pharmacology 5th. 北京大學醫學出版社. 2013: 26–36. ISBN 978-7-5659-0620-6. 引文格式1维护:显式使用等标签 (link)
^ The Nobel Prize in Physiology or Medicine 1994. Nobel Foundation. [2007-07-28]. (原始内容存档于2007-07-18).
^ Royal Swedish Academy of Sciences. The Nobel Prize in Chemistry 2012 Robert J. Lefkowitz, Brian K. Kobilka. 2012-10-10 [2012-10-10]. (原始内容存档于2012-10-12).
^ 8.08.18.28.38.48.58.6 David L. Nelson; Michael M. Cox; 等. Lehninger Priciples of Biochemistry 6th. New York: W. H. Freeman and Company. 2013: 436–484. ISBN 978-1-4292-3414-6. 引文格式1维护:显式使用等标签 (link)
^ Attwood TK, Findlay JB. Fingerprinting G-protein-coupled receptors. Protein Eng. 1994, 7 (2): 195–203. PMID 8170923. doi:10.1093/protein/7.2.195. (原始内容存档于2007-10-12).
^ Kolakowski LF Jr. GCRDb: a G-protein-coupled receptor database. Receptors Channels. 1994, 2 (1): 1–7. PMID 8081729.
^ Foord SM, Bonner TI, Neubig RR, Rosser EM, Pin JP, Davenport AP, Spedding M, Harmar AJ. International Union of Pharmacology. XLVI. G protein-coupled receptor list. Pharmacol Rev. 2005, 57 (2): 279–88. PMID 15914470. doi:10.1124/pr.57.2.5.
^ InterPro 互联网档案馆的存檔,存档日期2008-02-21.
^ Robinson DR, Wu YM, Lin SF. The protein tyrosine kinase family of the human genome. Oncogene. November 2000, 19 (49): 5548–57. PMID 11114734. doi:10.1038/sj.onc.1203957.
^ Ségaliny et al. (2015) "Receptor tyrosine kinases: Characterisation, mechanism of action and therapeutic interests for bone cancers". Bone Oncology 23;4 (1):1-12.[1] Retrieved on 2017-09-17
^ George J. Siegel; R. Wayne Albers. Basic neurochemistry: molecular, cellular, and medical aspects. Academic Press. 2006: 368– [2010-12-16]. ISBN 978-0-12-088397-4.
^ Gene Family: Ligand gated ion channels. HUGO Gene Nomenclature Committee. (原始内容存档于2017-11-14).
^ 道兰氏医学词典中的ligand-gated channel
^ Purves, Dale, George J. Augustine, David Fitzpatrick, William C. Hall, Anthony-Samuel LaMantia, James O. McNamara, and Leonard E. White. Neuroscience. 4th ed.. Sinauer Associates. 2008: 156–7. ISBN 978-0-87893-697-7.
^ Evans RM. The steroid and thyroid hormone receptor superfamily. Science. May 1988, 240 (4854): 889–95. PMID 3283939. doi:10.1126/science.3283939.
^ Olefsky JM. Nuclear receptor minireview series. The Journal of Biological Chemistry. October 2001, 276 (40): 36863–4. PMID 11459855. doi:10.1074/jbc.R100047200.
^ Lalli E, Sassone-Corsi P. Signal transduction and gene regulation: the nuclear response to cAMP. The Journal of Biological Chemistry. Jul 1994, 269 (26): 17359–62. PMID 8021233.
^ Rosen OM. After insulin binds. Science. Sep 1987, 237 (4821): 1452–8. Bibcode:1987Sci...237.1452R. PMID 2442814. doi:10.1126/science.2442814.
^ 23.023.123.2 李端 殷明. 药理学. 人民卫生出版社. 11-15. ISBN 978-7-117-08905-0
| ||||||||||||||||||||||||||||||||||||||||||||||||


