糖酵解
| 本条目部分链接不符合格式手冊規範。跨語言链接及章節標題等處的链接可能需要清理。(2015年12月13日) |

糖解作用总览
糖酵解(英语:glycolysis,又称糖解)是把葡萄糖(C6H12O6)转化成丙酮酸(CH3COCOO− + H+)的代谢途径。在这个过程中所释放的自由能被用于形成高能量化合物ATP和NADH。[1][2]
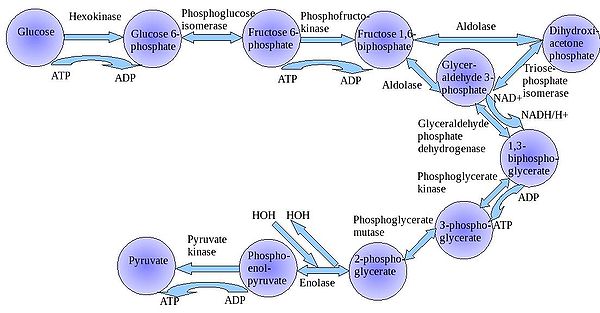
糖解作用是所有生物细胞糖代谢過程的第一步。糖解作用是一共有10个步骤酶促反应的确定序列。在该过程中,一分子葡萄糖会经过十步酶促反应转变成两分子丙酮酸(严格来说,应该是丙酮酸盐,即是丙酮酸的阴离子形式)。
糖解作用及其各种变化形式发生在几乎所有的生物中,无论是有氧和厌氧。糖酵解的广泛发生显示它是最古老的已知的代谢途径之一。[3]事实上,构成糖解作用及其并行途径的戊糖磷酸途径,在金属的催化下发生在还不存在酶的太古宙海洋。[4]糖解作用可能因此源于生命出现之前世界的化学约束。
糖解作用发生在大多数生物体中的细胞的胞质溶胶。最常见的和研究最彻底的糖解作用形式是双磷酸己糖降解途径(Embden-Meyerhof-Parnas途径,简称:EMP途径),这是被Gustav Embden,奥托·迈尔霍夫,和Jakub Karol Parnas所发现的。糖解作用也指的其他途径,例如,脱氧酮糖酸途径(Entner-Doudoroff途径)各种异型的和同型的发酵途径,糖解作用一词可以用来概括所有这些途径。但是,在此处的讨论却是局限于双磷酸己糖降解途径(EMP途径)。[5]
整个糖解作用途径可以分成两个阶段:[1]
- 准备阶段 – 在其中ATP被消耗,因此也被称为投入阶段。
- 放能阶段 – 在其中ATP被生产。
目录
1 糖解作用的总反应式
2 糖解作用的场所
3 糖酵解步骤
4 反应序列
4.1 准备阶段
4.2 放能階段
4.3 糖酵解中的不可逆反应
4.4 糖解作用中的调节位点
5 NADH的去路
6 能量转化
6.1 平衡点
6.2 无氧环境和有氧环境的能量转化
7 参看
8 参考文献
9 外部链接
糖解作用的总反应式
糖解作用的总体反应式:
- C6H12O6 + 2 NAD+ + 2 ADP + 2 H3PO4 → 2 NADH + 2 C3H4O3 + 2 ATP + 2 H2O + 2 H+
D-[葡萄糖] | [丙酮酸] | ||||
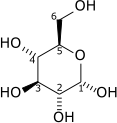 | + 2 [NAD]+ + 2 [ADP] + 2 [P]i | 2 |  | + 2 [NADH] + 2 H+ + 2 [ATP] + 2 H2O |
糖解作用
对于简单发酵,一个葡萄糖分子到两个丙酮酸分子的代谢具有净产率2个ATP分子。然后,大部分细胞将进行进一步的反应,以“偿还”用过的NAD+.,并产生最终产物乙醇或乳酸。许多细菌使用无机化合物作为氢的受体来再生NAD+。
细胞进行有氧呼吸合成更多的ATP,但不作为糖解作用的一部分。这些进一步的有氧反应使用来自糖解作用的丙酮酸和NADH + H+。真核生物有氧呼吸从每个葡萄糖分子产生额外的约34个ATP分子,但是其中大部分是由截然不同的机制,以在糖酵解的底物水平磷酸化产生的。
| 常见的单糖,包括糖酵解,糖异生,糖原和糖原分解的代谢。 |
|---|
 |
糖解作用的场所
糖解作用在细胞的细胞质中进行。早先人们只知道糖在无氧环境下可降解为乳酸,但今天人们终于清楚知道,不论有氧还是无氧环境,糖会经过同样的过程分解为丙酮酸。不同的则是在有氧条件下,丙酮酸被移出一分子的二氧化碳,剩餘的二碳以不穩定的鍵結連接至辅酶A(一種衍生自維生素B5的含硫化合物),形成具有異常活潑乙酰基(附著的乙酸鹽)的化學修飾物乙酰辅酶A,從而進入三羧酸循環。
在原核生物和真核生物的大部分缺氧细胞或组织(骨骼肌)中,丙酮酸会转化成乳酸,或者像糖类被酵母分解那样,转化为乙醇和二氧化碳(CO2)。在有氧环境下工作的组织(典型:心肌细胞)分解三碳的丙酮酸为乙酰辅酶A和二氧化碳,乙酰辅酶A会进一步行三羧酸循环分解为CO2和氢。氢会与氢载体烟酰胺腺嘌呤二核苷酸(NAD+)和黄素腺嘌呤二核苷酸(FAD)结合成(NADH和FADH2)。在线粒体裡进行的呼吸链,NADH和FADH的氧化会导致ATP的产生,能量会储存在ATP的高能磷酸键供细胞使用。
糖解作用是唯一一条现代生物都具有的代谢途径,出现时间很早。糖解作用最早可能发生在35亿年前第一个原核生物中。
糖酵解步骤
糖酵解的第一步是葡萄糖磷酸化为6-磷酸葡萄糖。不同细胞类型中所含有的酶也不一样,在所有的細胞中,皆有己糖激酶進行催化,而在肝和胰腺中,則另外含有一種稱為葡(萄)糖激酶(己糖激酶IV)的酵素[6]。磷酸化过程消耗一分子ATP,后面的过程证明,这是回报很丰厚的投资。细胞膜对葡萄糖通透,但对磷酸化产物6-磷酸葡萄糖不通透,后者在细胞内积聚并继续反应,将反应平衡向有利于葡萄糖吸收的那一面推移。之后6-磷酸葡萄糖会在磷酸己糖异构酶的催化下生成6-磷酸果糖。(在此果糖也可通过磷酸化进入糖解作用途径)
接着6-磷酸果糖会在磷酸果糖激酶的作用下被一分子ATP磷酸化生成1,6-二磷酸果糖,ATP则变为ADP。这里的能量消耗是值得的,:首先此步反应使得糖解作用不可逆地继续进行下去,另外,两个磷酸基团可以进一步在醛缩酶的参与下分解为磷酸二羟丙酮和3-磷酸甘油醛。磷酸二羟丙酮会在磷酸丙糖异构酶帮助下转化为3-磷酸甘油醛。两分子3-磷酸甘油醛会被NAD+和3-磷酸甘油醛脱氢酶(GAPDH)的氧化下生成1,3-二磷酸甘油酸。
下一步反应,1,3-二磷酸甘油酸转变为3-磷酸甘油酸。此反应由磷酸甘油酸激酶催化,高能磷酸键由1,3-二磷酸甘油酸转移到ADP上,生成两分子ATP。在此,糖解作用能量盈亏平衡。两分子ATP消耗了又重新生成。ATP的合成需要ADP作原料。如果细胞内ATP多(ADP则会少),反应会在此步暂停,直到有足够的ADP。这种反馈调节很重要,因为ATP就是不被使用,也会很快分解。反馈调节避免生产过量的ATP,节省了能量。磷酸甘油酸变位酶推动3-磷酸甘油酸生成2-磷酸甘油酸,最终成为磷酸烯醇式丙酮酸。磷酸烯醇式丙酮酸是高能化合物。最后,在丙酮酸激酶的作用下磷酸烯醇式丙酮酸生成一分子ATP和丙酮酸。此步反应也受ADP调节。
反应序列
糖解作用反应序列可被分为十个步骤
糖酵解代谢途径 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

准备阶段
通常视前五步为准备(或投入)阶段,因为这些步骤消耗能量以将葡萄糖转变为两个丙糖磷酸,即甘油醛-3-磷酸和磷酸二羟丙酮。
糖解作用準備階段的第一個步驟是將葡萄糖磷酸化,利用存於大部分動植物及微生物細胞內的己糖激酶所催化反應,此反应的标准自由能变化ΔG∘′=−4.0displaystyle Delta G^circ '=-4.0 千卡/莫耳,故這在胞內情況是不可逆的反應,將葡萄糖在6號碳處被ATP磷酸化,產生葡萄糖-6-磷酸。 千卡/莫耳,故這在胞內情況是不可逆的反應,將葡萄糖在6號碳處被ATP磷酸化,產生葡萄糖-6-磷酸。此步驟中的酵素,與己糖分子結合,本身的構形會發生改變,催化葡萄糖磷酸化,既然被命名為己糖激酶,代表其不僅僅催化右旋-葡萄糖,而也具備催化其他六碳的糖類,像是右旋-果糖以及右旋-甘露糖磷酸化的功能,而這種酶,以不同的同工酶存在於不同的生物體或組織中,在肌肉中,是一種調節性酶,當葡萄糖-6-磷酸和腺苷二磷酸的濃度超出正常值,便可以暫時的抑制己糖激酶,使之形成與利用的反應達到速率平衡。 而肝臟中,存在於一種特殊的己糖激酶名叫葡萄糖激酶,这种酶只對葡萄糖有專一性,並不會與所有己糖發生作用,而且此酶並不受葡萄糖-6-磷酸的抑制作用。它对葡萄糖的米氏常数为5~10毫摩尔/升,比己糖激酶的0.1毫摩尔/升要高很多,根据米氏常数的意义,当葡萄糖浓度很高时它才作用。由于肝脏是糖原的重要生成器官,因此当血糖浓度增高时,肝脏中的葡萄糖激酶就运作,生成了葡萄糖-6-磷酸,肝脏以此合成肝糖原。然而,此酶的合成受到胰岛素的诱导,因此,体内缺乏胰岛素的糖尿病患者的肝脏中此酶的合成速率低,影响肝脏中葡萄糖转变为葡萄糖-6-磷酸的速率,故分解葡萄糖以及合成肝糖原的过程受阻。 辅因子:镁离子 |
| ||||||||||||||||||||

| 糖解作用第二個步驟為將葡萄糖-6-磷酸轉化為果糖-6-磷酸,此步驟由磷酸葡萄糖異構酶所催化,此酶主要由高度純化自肌肉分離而出。此酶將前一步驟產物葡萄糖-6-磷酸的氧原子,由1號碳移至2號碳,將其異化為果糖-6-磷酸(F6P)。 此反应的标准自由能变化ΔG∘′=+0.4displaystyle Delta G^circ '=+0.4 |
| ||||||||||||||||||||


| 第三個步驟是將果糖-6-磷酸酸化為果糖-1,6-二磷酸,由磷酸果糖激酶所催化,這是糖解作用的第二個活化反應,將F6P的磷酸跟轉移到1號碳位置產生右旋-果糖-1,6-二磷酸。 此反应的标准自由能变化ΔG∘′=−3.40displaystyle Delta G^circ '=-3.40 辅因子:Mg2+ |
| ||||||||||||||||||||


| 前一步反应使得分子失稳,这使得己糖环可以被醛缩酶分成两个丙糖:二羟丙酮磷酸,一种酮,以及甘油醛-3-磷酸,一种醛。有两类醛缩酶:I类醛缩酶,存在于动物与植物中,以及II类醛缩酶,存在于真菌和细菌中;这两类醛缩酶使用不同的机制切断酮糖环。 |
| ||||||||||||||||||||||||||


磷酸丙糖异构酶迅速将二羟丙酮磷酸互变为甘油醛-3-磷酸,后者进入糖酵解的后续步骤。这是非常有用的,因为它引导二羟丙酮磷酸进入与甘油醛-3-磷酸相同的途径,简化了调控。 |
| ||||||||||||||||||||
放能階段
糖解作用的第二階段為放能階段,此階段的目的在於產生高能分子ATP和NADH。因為一個葡萄糖在準備階段時已經變成兩個丙糖,所以在放能階段中每個反應會發生兩次。最後產生2個NADH和4個ATP,使得單一葡萄糖在經過整個糖解作用後淨得2個NADH和2個ATP。ATP会用于其他需能反应,而NADH则会进入呼吸链或作为还原剂参与细胞内其他还原加氢反应。
| 两个丙糖分子被氧化并添加一分子无机磷酸,形成1,3-二磷酸甘油酸(1,3-BPG)。 被脱下的氢用于还原氢载体NAD+,形成NADH。 |
| ||||||||||||||||||||
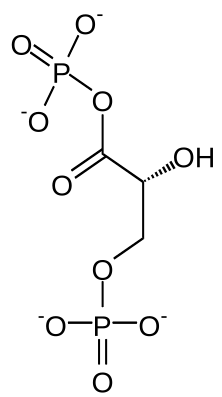
| 这步反应为磷酸甘油酸激酶将甘油酸-1,3-二磷酸的磷酸基团转移至ADP,形成甘油酸-3-磷酸和一份子ATP,在这一步,糖酵解过程达到了能量收支平衡:2分子ATP在先前的反应中被消耗,而在这步反应中有两分子ATP被合成。这步反应作为两步底物水平磷酸化中的一步,以ADP作为底物,所以当细胞ATP水平较高时,该步反应被抑制;因此该步反应也是糖酵解过程中重要的控速步骤之一。 |
| ||||||||||||||||||||

磷酸甘油酸变位酶催化形成甘油酸-2-磷酸。 |
| ||||||||||||||||||||

| 由烯醇化酶催化从甘油酸-2-磷酸转化为磷酸烯醇式丙酮酸。 辅基: |
| ||||||||||||||||||||

| 最后一步底物水平磷酸化由丙酮酸激酶催化,形成一份子丙酮酸和一分子ATP。 辅基: |
| ||||||||||||||||||||

糖酵解中的不可逆反应
人体可通过糖异生,即从非糖化合物,如丙酮酸和乳酸等物质重新合成葡萄糖。当肝或肾以丙酮酸为原料进行糖异生时,糖异生中的其中七步反应是糖酵解中的逆反应,它们有相同的酶催化。但是糖酵解中有三步反应,是不可逆反应。在糖异生时必须绕过这三步反应,代价是更多的能量消耗。
这三步反应都是强放能反应,它们分别是:
1葡萄糖经已糖激酶催化生成6磷酸葡萄糖ΔG= -33.5 kJ/mol
2 6磷酸果糖经磷酸果糖激酶催化生成1,6二磷酸果糖ΔG= -22.2 kJ/mol
3磷酸烯醇式丙酮酸经丙酮酸激酶生成丙酮酸ΔG= -16.7 kJ/mol
糖解作用中的调节位点
糖解作用在体内可被精确调节,这样一方面可以满足机体对能量的需要,另一方面又不会造成浪费。同时,当细胞内还进行糖异生的时候,调节就显得非常重要了,因为要避免空循环的发生。
调节是通过改变底物浓度,酶的活性实现的。
磷酸果糖激酶是其中最重要的限速酶,这也是巴斯德效应的关键参与者,它也决定了糖异生的速度,成为调节位点。AMP的浓度越高,酶的活性越高。就是当机体大量消耗了ATP,而相应又产生了很多AMP的时候,酶的活性提高,使得糖酵解按生成ATP的方向快速前进,以提高ATP产量。
NADH的去路
在细胞中,NADH与NAD+是处于动态平衡的。在糖酵解过程中生成的NADH必须被进一步氧化,转化为NAD+才能够让糖酵解持续进行。另外足够的NAD+是3磷酸甘油醛成为1,3二磷酸甘油酸这一步反应重要的前提。在此过程中NAD+会被还原为NADH+H+,即是氢载体,通过穿梭将氢带到呼吸链。[9]
NAD+的再生可通过这三种不同的过程来实现。
乳酸脱氢酶:由丙酮酸形成乳酸,此过程发生在骨骼肌及部分微生物中。
乙醇脱氢酶:经丙酮酸脱羧酶将丙酮酸转变为乙醛,再由乙醛经乙醇脱氢酶催化还原形成乙醇,此过程发生在大多数植物和一些产乙醇的微生物中。人类基因组可以编码乙醇脱氢酶,但是并没有编码丙酮酸脱羧酶的基因,因此无法将丙酮酸转化为乙醇。
线粒体穿梭:经线粒体穿梭途径进入线粒体的呼吸链生成ATP。
能量转化
平衡点
值得一提的是,生成1,6-二磷酸果糖后的大部分反应都是向能量升高的方向进行的,没有酶(磷酸果糖激酶(PFK),磷酸甘油酸激酶(PGK))的催化,是不会自发进行的。而糖酵解的逆过程--糖异生(从甘油等非糖物质生成葡萄糖)则容易进行,此过程用到大部分在糖酵解里面出现过的酶,除了提到的两位“车夫”外,它们只出现在糖酵解中。在糖异生这两步逆反应会放出大量的热,分别为-14及-24 kJ/mol。
无氧环境和有氧环境的能量转化

糖酵解中的NAD+和NADH+H+循环
在糖解作用中,每分子葡萄糖提供两分子ATP。真核生物的粒線体能同时从两分子丙酮酸中另外获得36分子ATP。能量转化的多少取决于在细胞质中产生的NADH + H+通过粒線体膜的方式。
不论在无氧还是有氧环境中,糖酵解成丙酮酸这一过程都能进行。3-磷酸甘油醛在3-磷酸甘油醛脱氢酶GAPDH的作用下脱氢。脱下的氢离子会将氧化剂(辅酶)NAD+还原成NADH + H+。NAD+会在呼吸链中再生。若在无氧环境,放热的(ΔGo´ = - 25 kJ/mol)乳糖脱氢酶(LDH)反应会再生NAD+:丙酮酸的还原会生成乳糖和再生NAD+(酵母则会使用另外两种酶—丙酮酸脱羧酶加乙醇脱氢酶)。下图可阐明此过程:
无氧环境下糖解作用GAPDH-和LDH-反应的相互联系,除了少部分NADH+H+会被磷酸甘油脱氢酶(GDH)转化外,大部分会用于再生NAD+。
参看
- 三羧酸循环
- 发酵
- 糖异生
- 磷酸戊糖途径
参考文献
^ 1.01.1 Glycolysis – Animation and Notes
^ Bailey, Regina. 10 Steps of Glycolysis.
^ Romano AH, Conway T. (1996) Evolution of carbohydrate metabolic pathways. Res Microbiol. 147(6–7):448–55 PMID 9084754
^ Keller, Ralser & Turchyn (2014). Non-enzymatic glycolysis and pentose phosphate pathway-like reactions in a plausible Archean ocean. Mol Syst Biol. 2014 Apr 25;10:725. doi: 10.1002/msb.20145228. PMID 24771084
^ Kim BH, Gadd GM. (2011) Bacterial Physiology and Metabolism, 3rd edition.
^ David L. Nelson & Michael M. cox. Lehinger. Principles of Biochemistry. 4th edition. Freeman. ISBN 0-7167-4339-6.
^ Reeves, R. E.; South D. J., Blytt H. J. and Warren L. G. Pyrophosphate: D-fructose 6-phosphate 1-phosphotransferase. A new enzyme with the glycolytic function 6-phosphate 1-phosphotransferase. J Biol Chem. 1974, 249 (24): 7737–7741. PMID 4372217. 引文使用过时参数coauthors (帮助)
^ Selig, M.; Xavier K. B., Santos H. and Schönheit P. Comparative analysis of Embden-Meyerhof and Entner-Doudoroff glycolytic pathways in hyperthermophilic archaea and the bacterium Thermotoga. Arch Microbiol. 1997, 167 (4): 217–232. PMID 9075622. 引文使用过时参数coauthors (帮助)
^ Stryer et al. Biochemistry, WH Freeman Complany, 2006. ISBN 0-71-678724-5
外部链接
(英文)糖酵解
(德文)Ulrich Helmich主页上的糖酵解
(德文)详细讲解糖酵解- NAD/NADH的结构
植物学在线上的糖酵解- 糖酵解基础
(英文)糖酵解中NADH和质子波动
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||

| ||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||
|


