真核生物
| 真核域 化石时期:中元古宙至今 冥古宙 太古宙 元古宙 显 | |
|---|---|
 | |
| 多种多样的真核生物 | |
科学分类 | |
| 总域: | 新壁總域 Neomura |
| 域: | 真核域 Eukaryota Whittaker & Margulis,1978 |
演化支 | |
| |
界 | |
真核生物(学名:Eukaryota)是其细胞具有细胞核的单细胞生物和多细胞生物的总称,它包括所有动物、植物、真菌和其他具有由膜包裹着的复杂亚细胞结构的生物。
真核生物与原核生物的根本性区别是前者的细胞内含有细胞核,因此以真核来命名这一类细胞。许多真核细胞中还含有其它细胞器,如粒線體、叶绿体、高尔基体等。
由于具有细胞核,因此真核细胞的细胞分裂过程与没有细胞核的原核生物也大不相同。
真核生物在进化上是单源性的,都属于三域系统中的真核生物域,另外两个域为同属于原核生物的细菌和古菌。但由于真核生物与古菌在一些生化性质和基因相关性上具有一定相似性,因此有时也将这两者共同归于新壁總域演化支。
科學家相信,從基因證據來看,真核生物是細菌與古菌的基因融合體,它是某種古菌與細菌共生,異種結合的產物。
目录
1 起源
2 真核生物的分类
2.1 传统分类
2.2 2005年国际原生生物学家协会分类
2.3 当前的争议与共识
3 真核细胞
3.1 内膜系统
3.2 内共生体
3.3 细胞骨架
4 繁殖
4.1 有性生殖
5 各种真核细胞
5.1 植物细胞
5.2 真菌细胞
5.3 动物细胞
6 参见
7 参考文献
起源
真核细胞之所以可以发展出有质膜包围的细胞核和细胞器,被认为有两个过程:膜内折和内共生。膜内折指始祖古核生物细胞的质膜内折成为真核细胞的膜系统。经过第二次内折,内质网膜形成高尔基体等细胞器。内共生学说指,古核生物吞入了需氧的、较小的、始祖原核生物(需氧真细菌,如变形细菌)和,由寄生过渡到共生,变为线粒体。通过类似的过程,古核生物细胞与较小的、始祖光合原核生物(如蓝细菌)共生,产生绿藻和红藻,第二次被古核生物吞入成为共生者,并演变为叶绿体。膜内折产生的原始的内质网膜系统,限制了吞入细菌的活动,避免了细胞成分被其“吃掉”。
真核生物的分类
传统分类
在古时候,人们就已认识到动物和植物应分为两类。在林奈建立分类阶元时,虽然存在一定疑惑但还是把真菌划为两界说中的植物界。而后来的学者把真菌独立成一界,即把真核生物分为四界:原生生物界、植物界、动物界、真菌界。显微镜发明后,所发现的许多原生生物根据是自养还是异养,划分为原生植物与原生动物,分别归入植物界与动物界。而海克尔增加了原生生物界,即把真核生物分为四界。
在20世纪80年代之前,单细胞的原生生物的分类还处于很混乱的阶段,直到DNA测序技术的成熟使系统发生学出现,改变了这一状况,也使人们开始重新谱写真核生物的分类。
2005年国际原生生物学家协会分类
根据系统发生学的研究,根据进化分支上的同源性,2005年的国际原生生物学家协会提出了一种在当时被普遍接受的真核生物分类方法,分作六类(supergroups)。这种分类方法并没有明确这些类是生物分类学上的哪一等级,但其它文献将其当作"界".
| 古虫界 (Excavata) | 鞭毛虫等原生生物 |
| 变形虫界 (Amoebozoa) | 阿米巴与黏菌等 |
| 后鞭毛生物 (Opisthokonta) | 动物、真菌、领鞭毛虫类 (choanoflagellate)等 |
| 有孔虫界 (Rhizaria) | 有孔虫、放射虫、某些变形虫原生生物 |
| 囊泡藻界 (Chromalveolata) | 不等鞭毛门 (Heterokonta)、定鞭藻门 (Haptophyta)、隐藻门 (Cryptophyta) , 囊泡虫总门 (Alveolata) |
| 泛植物界 (Archaeplastida) | 陆生植物、绿藻、红藻、灰胞藻 |
但是,这一"六界说"很快就遭到了新的研究的挑战,特别是对囊泡藻界是否有共同起源的质疑。真核生物的分类目前仍有很大的不确定性。例如,把真核生物分为单鞭毛生物(Unikonta)与双鞭毛生物(Bikonta),前者包括变形虫界与后鞭毛生物,后者其它四界。但是,很多研究表明单鞭毛生物与双鞭毛生物都不是同源的进化支。另外,有孔虫界应该并入囊泡藻界。泛植物界也应该不是同源的,需要被细分。
当前的争议与共识
一些研究拆分了囊泡藻界,并将其一部分与有孔虫界合并为SAR超类群,另一部分视为原始色素体生物的近缘类群 ,比如Burki等在2007提出的系统树
| |||||||||||||||||||||||||||||||||||||||||||
另一些研究则认为Hacrobia和古虫界的一部分应该属于原始色素体生物,和SAR超类群组成双鞭毛生物,变形虫和后鞭毛生物组成单鞭毛生物:
| ||||||||||||||||||||||||||||||||||||||||
还有一些研究认为囊泡藻界不应被拆分,而是将SAR超类群归入扩大的囊泡藻界,如Rogozin 等在2009年的一项研究:
| |||||||||||||||||||||||||||||||||||||
许多观点将扩大的囊泡藻界视为原始色素体生物的近缘类群:
| |||||||||||||||||||||||||||||||||||||
2009年一项研究详细研究了已有的真核生物分类的证据,提出了如下分类:
| 真核生物 |
| |||||||||||||||||||||||||||||||||
2012年的一项研究也支持单鞭毛生物与双鞭毛生物的划分:
| 真核生物 |
| ||||||||||||||||||||||||||||||
现在广泛认为有孔虫和不等鞭毛类及囊泡虫类是姊妹类群,同属于一个超类群,因此有孔虫也不再视为真核生物的主要类群之一,同时单鞭毛生物及其子类群变形虫和后鞭毛生物都被视为单系群,即动物和真菌同属一个单系群--后鞭毛生物。除此之外,还没有其它共识。
真核细胞

真核生物的內膜系统
真核细胞体积通常远大于原核细胞,其内部有各种内膜结构构成的内膜系统。细胞器与微丝、微管、中间丝等细胞骨架在细胞的功能和结构上起到了重要的作用。真核DNA线性地划分在几个染色体上,平时以染色质形态存在于细胞核中,在有丝分裂时由微管组成的纺锤丝分离。
内膜系统
真核细胞包含多种膜结构,统称为内膜系统。膜可以部分脱离,产出独立的小室,称为小泡或液泡。很多细胞通过胞吞作用摄入食物和其他物质,外层膜首先入鞘,然后收紧形成独立的小泡弹出。大多其他由膜包围的细胞器都可能形成自这种液泡。
细胞核由两层膜包围,常称为核膜,包含核孔,使得物质能够进出。核膜的管状、片状突出形成了内质网,负责蛋白质的生成和转运。粗面内质网上有核糖体,进行蛋白质合成,这些蛋白质随后进入细胞内腔,再进入小泡中,然后从光面内质网弹出。对于大多真核细胞,这些携带蛋白质的小泡释放到一堆扁平囊泡中,即高尔基体,并进一步改装。
小泡可以被特化,用于多种不同目的。例如,溶酶体装有可以分解食物的酶,過氧化物酶用于分解有毒的過氧化物。很多原生生物有可收缩的液泡,用于收集和排出多余的水,排出小体可以喷出物质,用于击偏捕食者,或抓住猎物。而在高等植物中,中央液泡占据细胞多半的体积,该液泡也维持了它的渗透压。

粒线体结构: 1:内膜 2:外膜 3:线粒体嵴 4:基质
内共生体
粒线体是几乎所有真核细胞共有的细胞器,有两层膜,即两层磷脂双分子层所包裹,内膜向内折叠成嵴,在此进行有氧呼吸。线粒体有自己的DNA,今天通常认为是由内共生学说的原核生物发展而来,很可能是接近立克次體的變形菌門細菌。少数原生生物中没有线粒体,却发现具有由线粒体起源的细胞器,如氢化酶体和纺锤剩体,因此很可能这些原生生物是在之后才丢失了线粒体。
植物和很多藻类拥有细胞器色素体。同样的,这些色素体也有自己的DNA,起源于内共生,这次的内共生体是藍菌門细菌。色素体又称质体,最常见的就是叶绿体,像蓝菌门细菌一样也有叶绿素,通过光合作用合成有机物(如葡萄糖)。其他色素体参与储存食料。虽然色素体很可能是单起源的,但并非所有含有色素体的细胞也是单起源的,实际就有些真核细胞通过多重内共生或摄入获得色素体。
内共生起源也许能适用于细胞核,此前鞭毛也被认为起源于螺旋體。但由于缺乏细胞生物学证据,且难以与细胞繁殖过程调和,螺旋體一说未被广泛接受。
细胞骨架
许多真核生物具有细长且能运动的突起,称为鞭毛,以及较短但类似的结构—纤毛,合称为波动足,与真核生物的运动、感觉和摄食密切相关,主要成分是微管蛋白。这些细胞器与原生生物的鞭毛具有鲜明的不同,基体伸出一簇微管,支撑起它们。基体也被称作毛基体或中心粒,是中心体的主要组成部分。鞭毛本身也可以再有茸毛,以及鳞片,连接膜和内杆,它们又与细胞质相连。
微丝结构由肌动蛋白和结合了肌动蛋白的蛋白质构成,例如α-肌动蛋白、丝束蛋白和细丝蛋白也存在于膜下皮质层和皮质束众。微管中的马达蛋白,如动力蛋白,或驱动蛋白和肌动蛋白,如动力蛋白,为系统提供了动态特性。
即使是没有鞭毛的细胞也常有中心粒,但开花植物和松柏门植物则没有。中心粒通常成组出现,称作动胞器(kinetids),形成多种不同的微管根,它们构成了细胞骨架结构的主要成分,往往在多次细胞分裂的过程中合成,一个鞭毛从亲代保留下来,另一个则据亲代衍生而来。此外,中心粒也可能与细胞核分裂时纺锤体的形成有关。
繁殖
以有丝分裂的形式,细胞核的分裂通常与细胞分裂得以相互协调,这一过程使得每一个子代细胞核都得到亲代的一条染色体拷贝。在大多真核细胞中,还有另一种有性繁殖过程,即减数分裂,这一过程中,二倍体亲代细胞经由两次分裂成为单倍体,DNA的数量减半。然而,减数分裂本身也分很多种类。[1]
相比原核细胞,真核细胞有更小的表面积-体积比,因此新陈代谢速率较慢,细胞周期更长。在有些多细胞生物体中,一些专门用于新陈代谢的细胞会扩大表面积,如小肠绒毛细胞。
有性生殖
现在的真核生物广泛地采用有性生殖,并有证据显示,这是真核生物原始、根本的特性。基于系统发生学分析,生物学家达克斯和罗杰提出,真核细胞共有的祖先进行随机性行为。减数分裂中核心的一组基因出现在阴道毛滴虫和肠贾第虫中,而这两种生物此前被视为是无性的。在真核细胞进化树上,这两个物种很早便分离出来,因此可以推断,减数分裂的核心基因存在于所有真核生物的那个共同祖先中,也因此这一祖先是有性的。其他对真核物种的研究也揭露了生殖周期的证据。例如寄生原生生物利什曼原蟲近来显示存在生殖周期。证据显示,此前被认为是无性的阿米巴虫,在古代也是有性的,并且如今大多无性生物也是近期才独立地进化为无性的。
各种真核细胞
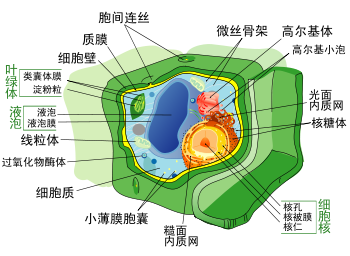
典型的植物细胞结构
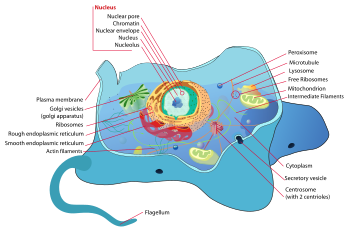
典型的动物细胞结构
植物细胞
植物细胞也与其它真核细胞有很多不同。其特征是:
- 由单层膜所包围的中央大液泡,维持细胞渗透压,其中的液体也是植物汁液的主要来源。
纤维素、半纤维素与果胶构成的细胞壁,保护原生质体。这与含有几丁质成分的真菌、粘菌和水霉细胞壁不同。
胞间连丝通过纹孔穿过细胞壁使每个植物细胞与其相邻细胞连接,这与功能类似的动物细胞间隙连接系统有所不同。- 含有许多两层膜的色素体(即原始色素体,植物也是原始色素体生物中的一类),特别是富含叶绿素的叶绿体,进行光合作用并使植物呈绿色。
- 高等植物,包括松柏和开花植物(被子植物)缺少鞭毛和中心粒。

真菌菌丝细胞 1:菌丝壁 2:隔膜 3:线粒体 4:液泡 5:麦角固醇晶体 6:核糖体 7:核 8:内质网 9:脂质体 10:质膜 11:顶体 12:高尔基体
真菌细胞
真菌细胞与动物细胞最为相似,但存在以下不同:
- 拥有甲壳素组成的细胞壁。
- 难以区分单个细胞:高等真菌的菌丝拥有隔膜,但隔膜可以允许细胞质中的细胞器甚至是细胞核通过;原始真菌的菌丝很少或根本没有隔膜,所以每个生物体本质上是一个巨大的多核超级单体。
- 只有最原始的真菌壶菌门拥有鞭毛。
动物细胞
动物细胞是组成动物组织的基本单元。动物细胞与其它真核细胞有巨大差异,主要体现在缺少细胞壁、大液泡等,但具有能进行胞吞、胞吐的小液泡。也正是因为缺乏细胞壁,动物细胞可以转换成不同的形态,吞噬細胞正是具有这点才能进行吞噬作用。
动物细胞可以分化成许多种类型,比如成人体内大约有210种细胞。
参见
- 已测序真核生物基因组列表
- 有性生殖的演化
- 三域系統
- 内共生
参考文献
^ 吴相钰; 陈守良; 葛明德. 陈阅增普通生物学 4. 高等教育出版社. 2014. ISBN 9787040396317.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


